
Celebrating an unsung hero of genomics: how Albrecht Kossel saved bioinformatics from a world of hurt
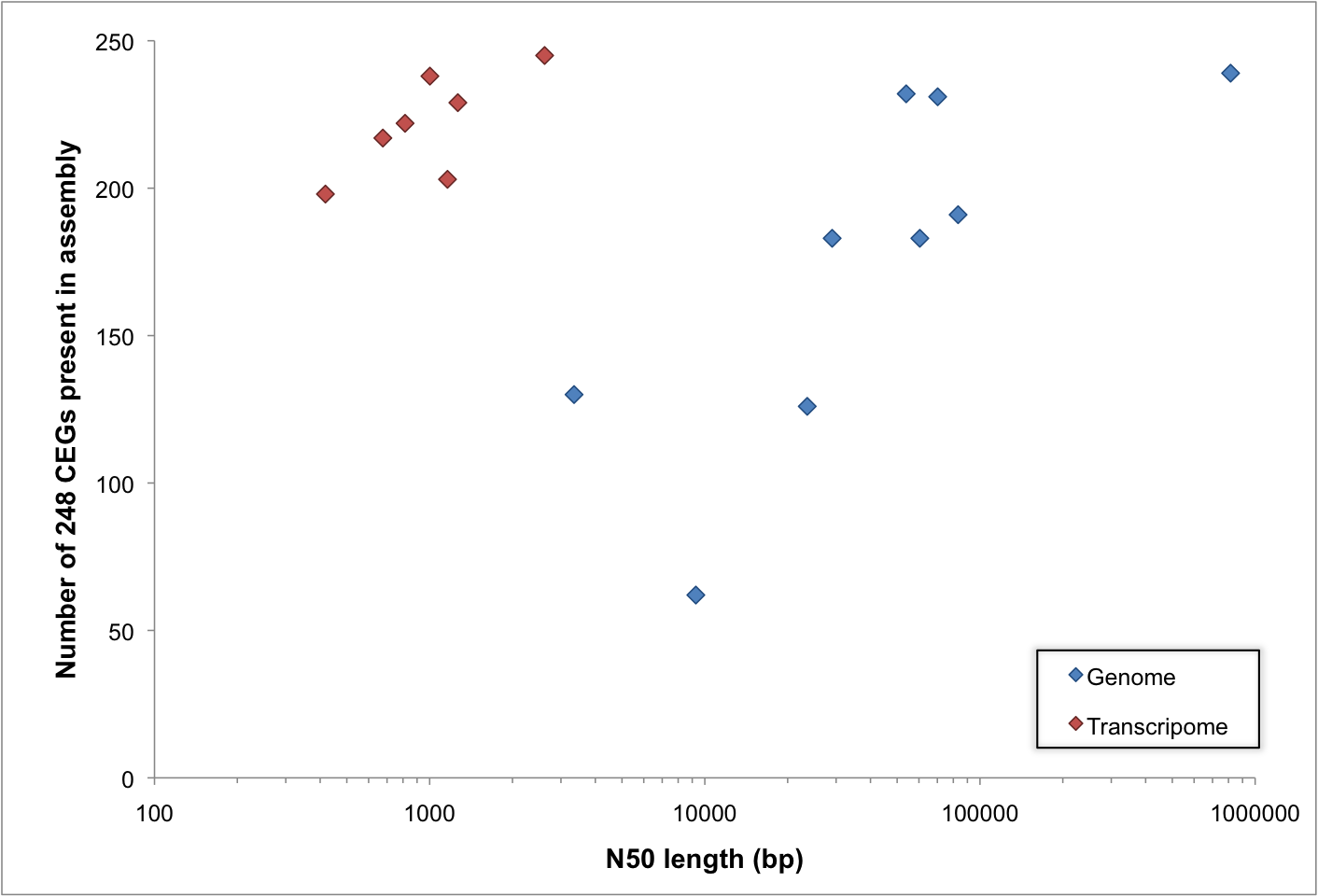
The following image is from one of the first publications to ever depict a DNA sequence in a textual manner:

From Wu & Kaiser, Journal of Molecular Biology, 1968.
This is taken from the 1968 paper by Wu and Kaiser: Structure and base sequence in the cohesive ends of bacteriophage lambda DNA. For almost half a century since this publication, it has become the norm to simply represent the sequence of any DNA molecule as a string of characters, one character per base. But have you ever stopped to consider why these bases have the names that they do?
Most of the work to isolate and describe the purines and pyrimidines that comprise nucleic acids came from the work of the German biochemist Albrecht Kossel. Between 1885 and 1901 he characterized the principle nucleobases that comprise the nucleic acids: adenine, cytosine, guanine, and thymine (though guanine had first been isolated in 1844 by Heinrich Gustav Magnus). The fifth nucleobase (uracil) was discovered by Alberto Ascoli, a student of Kossel.
Kossel's work would later be recognized with the 1910 Nobel Prize for Medicine. It should be noted that Kossel didn't just help isolate and describe these bases, he was also chiefly responsible for most of their names.

Albrecht Kossel, Image from wikimedia
Guanine had already been named based on where it had first been discovered (the excrement of seabirds known as guano). Adenine was so named by Kossel because it was isolated from the pancreas gland ('adenas' in Greek). Thymine was named because it was isolated in nucleic acids from the thymus of a calf. Cytosine — the last of the four DNA bases to be characterized — was also discovered from hydrolysis of the calf thymus. Its name comes from the original name in German ('cytosin') and simply refers to to the Greek prefix for cell ('cyto').
While this last naming choice might seem a little dull, all bioinformaticians owe a huge debt of thanks to Albrecht Kossel. Thankfully, he ensured that all DNA bases have names that start with different letters. This greatly facilitates their representation in silico. Imagine if he had — in a fit of vanity — instead chosen to name these last two bases that he characterized after himself and his daughter Gertrude. If that had been the case then maybe we would today be talking about the bases adenine, albrechtine, guanine, and gertrudine. Not an insurmountable problem to represent with single characters — we already deal with the minor headache of representing purines and pyrimidines differently (using R and Y respectively) — but frankly, it would be a royal pain in the ass.
Thank you Albrecht. The world of bioinformatics is in your debt.











{kind=link}